UNIVERSITY of EDINBURGH

DEPARTMENT of AGRICULTURE
HONOURS THESIS
Title Factors affecting the Germination and Establishment of Monogerm Sugar Beet.
Author's Name Robert A R Meikle
Degree B.Sc.
Date May 1981
FACTORS AFFECTING THE
GERMINATION AND
ESTABLISHMENT OF MONOGERM SUGAR BEET
by
ROBERT ANDREW ROSS MEIKLE
in partial fulfilment of the
requirements
for the Degree of BSc in
Agriculture
with Honours in Crop
Production Science.
at the
School of Agriculture
University of Edinburgh
May 1981
ACKNOWLEDGEMENTS
I should like to express my
sincere thanks to Dr Fred Harper for his guidance and much appreciated
criticism throughout the preparation of this thesis. I would also like to thank the staff at the crop production glass-houses
for their help with the experiments.
Thanks are also due to Mrs E Grosse and Typerite for their excellent
typing of the script.
CONTENTS
Page
|
|
LIST OF FIGURES |
|
|
(i) |
||||
|
1. |
INTRODUCTION |
|
|
1 |
||||
|
2. |
REVIEW OF LITERATURE |
|
|
3 |
||||
|
|
2.1 |
Introduction to Review |
|
3 |
||||
|
|
2.2 |
Seed production and seed treatments |
|
3 |
||||
|
|
|
2.2.1 |
Introduction |
|
3 |
|||
|
|
|
2.2.2 |
A brief history of
monogerm seed |
|
4 |
|||
|
|
|
2.2.3 |
Sugar beet breeding |
|
4 |
|||
|
|
|
2.2.4 |
Varieties |
|
5 |
|||
|
|
|
2.2.5 |
Commercial seed production |
|
5 |
|||
|
|
|
2.2.6 |
Post harvest treatments |
|
6 |
|||
|
|
|
2.2.7 |
Pelleting |
|
7 |
|||
|
|
|
2.2.8 |
Alternative seed
treatments |
|
8 |
|||
|
|
2.3 |
Factors affecting germination |
|
12 |
||||
|
|
|
2.3.1 |
Introduction |
|
12 |
|||
|
|
|
2.3.2 |
Seed storage |
|
12 |
|||
|
|
|
2.3.3 |
Germination tests |
|
13 |
|||
|
|
|
2.3.4 |
Effects of inhibitors on
germination |
|
14 |
|||
|
|
|
2.3.5 |
The germination process and the involvement of air
and water |
|
15 |
|||
|
|
2.4 |
Factors affecting the establishment of beet |
|
19 |
||||
|
|
|
2.4.1 |
Introduction |
|
19 |
|||
|
|
|
2.4.2 |
Plant population |
|
20 |
|||
|
|
|
2.4.3 |
Cultivation and seed bed
preparations |
|
21 |
|||
|
|
|
2.4.4 |
Drilling |
|
21 |
|||
|
|
|
2.4.5 |
Field emergence |
|
23 |
|||
|
|
|
2.4.6 |
Predicting sugar beet
emergence |
|
24 |
|||
|
|
|
2.4.7 |
The effect of seed size on
emergence |
|
26 |
|||
|
|
|
2.4.8 |
Other emergence factors |
|
27 |
|||
CONTENTS (Contd)
Page
|
|
2.5 |
Alternative methods of establishing beet |
|
29 |
||||||
|
|
|
2.5.1 |
Transplanting |
|
29 |
|||||
|
|
|
2.5.2 |
Autumn sowing |
|
30 |
|||||
|
|
|
2.5.3 |
Conclusion |
|
30 |
|||||
|
3. |
EXPERIMENTS |
|
|
31 |
||||||
|
|
3.1 |
Materials and Methods |
|
31 |
||||||
|
|
|
3.1.1 |
Description of seed lots |
|
31 |
|||||
|
|
|
3.1.2 |
Seed characteristics |
|
31 |
|||||
|
|
|
3.1.3 |
Germination and emergence
tests |
|
32 |
|||||
|
|
|
3.1.4 |
The effect of
water-soluble inhibitors on cress seed germination |
|
36 |
|||||
|
|
3.2 |
Results |
|
36 |
||||||
|
|
|
3.2.1 |
Seed characteristics |
|
36 |
|||||
|
|
|
3.2.2 |
The germination
experiments |
|
38 |
|||||
|
|
|
3.2.3 |
The emergence test |
|
57 |
|||||
|
|
|
3.2.4 |
The effect of
water-soluble seed extracts on cress seed germination |
|
57 |
|||||
|
|
3.3 |
Discussion |
|
60 |
||||||
|
|
|
3.3.1 |
Germination tests at standard temperatures and
7.5°C with untreated seed |
|
60 |
|||||
|
|
|
3.3.2 |
Seed characteristics |
|
63 |
|||||
|
|
|
3.3.3 |
Germination tests with treated
seed |
|
64 |
|||||
|
|
|
3.3.4 |
The emergence test |
|
69 |
|||||
|
|
|
3.3.5 |
Conclusion |
|
70 |
|||||
|
4. |
GENERAL DISCUSSION: THE POTENTIAL OF SEED
TREATMENTS TO IMPROVE FIELD EMERGENCE |
|
|
71 |
||||||
|
5. |
SUMMARY |
|
|
76 |
||||||
|
6. |
CONCLUSIONS |
|
|
77 |
||||||
|
7. |
REFERENCES |
|
|
78 |
||||||
LIST OF FIGURES
Page
1. The relationship between seed weight and
true seed weight for Seed Lot 1 39
2. The relationship between seed weight and
true seed weight for Seed Lot 2 40
3. The relationship between seed weight and
true seed weight for Seed Lot 3 41
4. The relationship between seed weight and
true seed weight for Seed Lot 4 42
5. The relationship between seed weight and
true seed weight for Seed Lot 5 43
6. Germination pattern of five seed lots under
standard germination test
conditions 47
7. Germination pattern of five seed lots under
low temperature conditions
(constant 7.5°C) 49
8. Germination pattern of three seed lots (advanced in water)
under standard germination test conditions
52
9. Germination pattern of five seed lots (advanced
in water) under low
temperature conditions 53
10. Germination pattern of five seed lots (with GA3,
solution in place of distilled H20 in the petri-dishes) otherwise
under standard
germination test conditions 55
11. Germination pattern of three seed lots (advanced in GA3 solution)
under standard germination
conditions
56
1. INTRODUCTION
The final yield of sugar
obtained from the sugar beet crop (Beta vulgaris L.) is determined by many factors, including: the length of the growing
season, the incidence of diseases and pests, and the prevailing weather
conditions (Biscoe, Draycott & Jaggard 1980). These factors have a direct or indirect effect on photosynthesis
and consequently the amount of assimilate translocated to the roots. It is
therefore the duration of the leaves which can produce assimilate (i.e.
excluding excessively shaded and diseased leaves) that determines the maximum
potential yield.
The actual yield obtained is
determined by the amount of solar radiation intercepted and utilised by the
leaf canopy over the - whole growing season.
Scott, English, Wood & Unsworth (1973) have shown that a good linear
relationship between sugar yield and both total incident and intercepted
radiation exists.
The grower cannot of course
control the amount of incident radiation but his objective must be to ensure as
much of it is intercepted as possible.
However under the ambient Spring soil temperatures of the U.K., sugar
beet is very slow in emerging and establishing, and complete leaf coverage does
not occur until July.
It was estimated by Scott
& Jaggard (1978) that just over 50% of the total radiation is intercepted
by crops in the ground between April and October. The rest strikes bare ground and is wasted.
There is clearly
considerable scope for improving the efficiency of interception by achieving
full leaf coverage sooner It is in fact the slow rate of growth over the early
part of the season that results in late leaf coverage and poor
-1-
-2-
radiation interception. Growers would welcome any feasible method of improving the early
growth of the crop.
The objective of this thesis
is to discuss the early stages of sugar beet growth and the various problems
affecting establishment that exist with sugar beet seed, both in its production
and performance in the field.
-3-
2. REVIEW OF LITERATURE
2.1 Introduction to review
The literature reviewed in
this thesis considers three main areas, namely seed production, seed
germination and field establishment.
Standard methods and various alternative procedures for treating and
testing beet seeds are firstly described and discussed and then related to
field performance. It will be explained
why establishment is still a problem in modern crops and why seed quality is of
major importance.
In the review and thereafter
whole fruits will be referred to as seeds, embryos as true seeds, and the rest
of the fruit as the seed coat and the ovary cap (which is considered to be part
of the seed coat), as the seed cap unless otherwise indicated.
2.2 Seed production and seed
treatment
2.2.1 Introduction
This section is mainly
concerned with monogerm sugar beet seed as multigerm seeds are no longer used
for sugar production.
The considerable
difficulties encountered in breeding and producing good quality monogerm seed
are outlined and modern varieties described.
As freshly harvested seed is
of poor quality and cannot be used successfully in practice because it contains
both empty seeds and multigerms the various processing procedures and
alternatives are described in detail.
The limitations of certain treatments are considered, as perfect seed
lots cannot be produced on a commercial scale.
-4-
2.2.2 A brief history
of monogerm seed
The advantages of seeds
containing only one true seed were first realised in the early 1900's, but
early Breeding Projects in the U.S.A. failed to produce plants with more than
75% single flowers. The idea was
abandoned until the 1930's when it was re-investigated in Russia. Plants with complete monogermity were
produced but other growth features were poor and commercial varieties were not
available until the 1960's due to severe breeding difficulties (Orlovskii 1957)
& (Savitsky 1952). Today however,
monogerm varieties have almost completely replaced multigerms.
2.2.3 Sugar beet breeding
Breeding sugar beet is
complicated by its biennial habit which makes the period taken to produce new
varieties much longer than with most other species. It is also an open or out pollinating crop which makes uniformity
difficult to achieve. However, several
methods of breeding can be used, e.g. mass selection where the largest and best
shaped roots are selected to produce seeds which are planted and the best
progeny selected for further seed production.
The method can be used to reliably improve existing varieties when
crossed with other plants with desirable characters. Other methods used singly or in various combinations include
hybridization polyploidy and male sterile lines. Some of the main objectives in sugar beet breeding are:-
1
The
monogerm character for commercial crops but multigerm pollinators are still
necessary.
2
High
sugar yield per ha as the best compromise between root yield and sugar %.
3
Low
bolting tendency.
4
Good
germination and early growth.
-5-
5
High
purity.
6
Good
root shape.
7
Disease
resistance to virus yellows & downy mildew.
(Ellerton 1978).
2.2.4 Varieties
All nine varieties on the
recommended list are genetic monogerm and produce over 90% single plants after
processing (NIAB 1980). All modern
varieties produce similar yields in trials, and have sugar contents over 17%. There are varietal differences in downy
mildew resistance, bolting susceptibility, size of tops, and field
emergence. Nomo and the newly
recommended Monoire show low bolting tendencies when sown early, while Sharp's
Klein Monobeet is poor in this respect.
Vytomo and Salohill are much less resistant to downy mildew than Monoire
or Bush Mono "G".
2.2.5 Commercial seed
production
Sugar beet seed is usually
grown away from root crops because of downy mildew and virus yellows carry over
and possible pollen contamination from bolters (Scott & Longden 1973). Most of the seed used for root crops is
triploid hybrid monogerm which is produced by mixing and then sowing together
an inbred diploid male sterile monogerm, and a tetraploid multigerm to act as a
pollinator. The mixed progeny is mechanically separable into triploid Monogerm
and multigerm seed (Johnson 1980, personal communication).
When seed crops are in
flower problems such as pollen contamination from weed beet, wild relatives or
low fertilisation overall due to unsuitable weather can occur, but the amount
of viable seed produced can be low for a number of reasons even if
-6-
good fertilisation is achieved (Scott & Longden
1973), (Battle & Whittington 1969a).
The maturity of the fruit at harvest can affect seed performance. More mature fruits have better developed
embryos and a lower concentration of germination inhibitors (Scott &
Longden 1973) and should therefore give better establishment of the root crop.
The actual numbers maturing
at harvest depends on the season and the date harvested. Wet seasons delay rate of maturing and often
seed is harvested early to reduce losses by windshake. Tekrony & Hardin
(1969) looked for the location of under-developed seed on seed plants and found
them on all parts but their occurrence was more frequent on tertiary lateral
branches. Field removal is therefore
not possible and it is also difficult to do so in processing unless there is a
relationship between "seed" size and under-developed seeds.
2.2.6 Post harvest
treatments
The standard post harvest
treatment to raw seed is a complicated and exacting operation but in outline
the following is carried out: -
1)
Pre-cleaned
(to remove stick & leaf) and dried to below 15% moisture at a temperature
not greater than 120°F (49°C).
2)
Cleaned
with aspiration using round hole screens to remove small and dead seeds.
3)
Monogerm
seed is separated from multigerm seed using a combination of round hole and
slotted screens.
4)
The
monogerm seed extracted is graded to size specifications - usually 2.75 - 4.00
mm.
5)
The
graded monogerm seed is gravity separated to extract light "weak"
seeds to improve germination.
-7-
6)
The
product is soaked in a fungicide solution of organic Ethyl Mercury Phosphate
(E.M.P.) for twenty minutes - by using a concentration of 40 ppm. E.M.P. and a ratio weight of three parts
water to one part seed.
7)
The
final seed product is then regraded to size specifications (Lindsay 1980,
personal communication).
8)
Pelleted.
2.2.7 Pelleting
Nearly all the seed sown in
the U.K. is pelleted as there are several advantages over raw-rubbed seed. The major advantage is more accurate
drilling and others are safe dressing with fungicides, protection from mechanical
damage and sowing depths and spacings are easily checked (Charlesworth 1978).
There are many materials and
methods which can be used for Clays such as Cellite, Montmorillonite,
Vermiculite or Bentonite, or other materials such as cork, peat, chalk, sand or
even beet cortex can be used. Non-clay
materials may need a sticker such as ethyl cellulose to hold the pellet
together. Clay structures generally
adhere with water only, but they need sufficient physical strength to resist
damage in the drill but breakdown easily on contact with soil water.
The three main methods of
pelleting are stamping, coating or rolling.
Stamping is a dry process and is therefore rapid but any additive must
be evenly distributed throughout the pelleting material, while coating and
rolling permit layering of additives anywhere within the pellet.
Drying after pelleting is
necessary and slows the procedure down (Longden 1975).
-8-
The most popular process in
the U.K. is the Germain's "Filcoat".
About 1600 tonnes of seed are pelleted annually with a secret clay-based
medium (Charlesworth 1978), plus methiocarb insecticide @ 4 Kg/1000 Kg seed
(Johnson 1980, personal communication). Manganese Oxide can also be
incorporated for use in deficient soils (Farley & Draycott 1978).
2.2.8 Alternative seed treatments
Sugar beet seed has to be
processed between harvesting and sowing to eliminate inferior seed, and give
good seed a better chance of good and fast emergence. The standard procedure listed removes empty and shrivelled seeds
and multigerms, and gives protection against some pests. and pathogens but
often field performance is far from perfect, therefore better treatments should
be sought. The problem in finding
suitable alternatives is not that treatments do not improve seed performance
when used singly but more often because there are practical difficulties or
incompatibility with existing treatments or simply the cost and time involved
(Tonkin 1979).
(i) Washing
Longden (1973) showed how 21
rinses in 3.5 hours gave the best response in glasshouse and field trials with
natural and partially processed seed; but when agitated, washed seeds showed a
greater tendency to lose seed caps and therefore this was an unsuitable
treatment. Washing had similar effects
on the performance of both mature and immature seed.
(ii) Advancing
The object of advancing is
to allow seeds to imbibe water and develop physiologically up to the point of
-9-
radicle elongation and then
be dried back, holding the seeds at this point before sowing. This reduces the period between sowing and
emergence in the field.
The optimum advancing
technique for sugar beet was determined by Longden (1971), (equal weights of
seed and water in airtight containers for 24h, repeated 3 more times with a 48h
air drying interval after each soaking).
Advancing increases the number of cells in the true seed but the
treatment does not increase the size of individual cells so that the
germination process does not therefore result in large changes in embryo volume
before the radicle elongates, but like washing, advancing increases the
tendency for premature loss of the seed cap.
An extremely gentle drill mechanism would therefore be required. Fluid drills may be developed for practical
use.
While washing was calculated
to be equivalent to a very much higher than normal rainfall over the seed
plant, rainfall effects are more similar to advancing than washing, i.e. a
small amount of water in contact with the seed for a longer time.
(iii)
Osmotic priming
This treatment is also
designed to get the seed in a better physiological state for quick
emergence. Trials with various
concentrations and priming periods in polyethylene glycol or salt (KNO3 +
K3 P04 H20), (Longden, Johnson, Darby &
Salter 1979) gave poor and inconsistent results and no reduction in the
-10-
time spread of
germination. Priming is therefore too
unreliable to use in practice.
(iv) Water steeping
Longden et al (1979)
also looked at the effects of a 24h soak in 10 x the seed volume of water @ 1°C followed by re-drying. Steeping gave uniform germination but poor
field performance. Scott, Wood &
Harper (1972) however found that a 24h soak in water @ 20°C hastened emergence
and improved final germination. The
steeping process probably has effects on physiological developments and also
removes germination inhibitors.
(v) Plant growth regulators
Scott et al (1972)
compared steeping seed in solutions of kinetin (K), 6-benzyl-aminopurine (BA)
and gibberellic acid (GA3) with water only. All solutions gave better field emergence
than water but also showed a response to concentration. The optimum concentrations were BA 1 - 10
ppm, K 50 - 100 ppm, GA3 - 100 ppm which nearly doubled the seedling
weights.
(vi) Size grading of seed
Longden, Scott & Wood
(1974) devised a size grading method which could produce a seed lot containing
90% monogerms and a germination of 80%, providing the unprocessed raw seed had
at least 50% germination. The method
included grading by thickness to remove multigerms, and grading by diameter and
aspiration to remove small and non-viable seeds. The variation in size range was also reduced in the process. However, an ideal sequence for size grading
cannot be
-11-
formulated as there is much
variation between original seed lots due to effects of season, seed production
agronomy and variety. The optimal
adjustment and sequence for one seed lot may be totally unsuitable for another.
(viii) Treatment combinations
Any number of treatments can
be used on a seed lot but to be of value they should be
a) fully compatible,
b) additive in effect or, preferably,
c)
synergistic
(Longden 1976).
Longden (1976) tried
washing, E.M.P. steeping, Thiram soaking and advancing in sequence on a seed
sample of Amono graded 3.18 - 3.57 mm.
One or more of the treatments were omitted on other Amono samples so that a total of 16 different
combinations of treatments were obtained for comparison. When analysed, different aspects of seed
performance were affected and complex interactions occurred - some combinations
gave faster emergence, others larger shoots.
In general, as more treatments were applied detrimental effects were
observed, e.g. E.M.P. had positive effects on seed performance when used
singly, but negative in combination, (when additional soaking let Hg penetrate
the embryo). It was probable that there
was considerable overlap in the effects of single treatments and there was no
evidence of cumulative or synergistic benefits from combinations. Therefore only one treatment involving water
or a solution should be used on a seed lot and in practice this is the E.M.P.
steep to control seed borne fungi.
-12-
2.3 Factors affecting germination
2.3.1 Introduction
This section is mainly
concerned with laboratory seed testing as affected by storage, natural
inhibitors and various treatments. The
most important factors in a petri-dish test are the access of oxygen to the
true seed and of water which is discussed in some detail. The importance and limitations of
germination tests are considered.
2.3.2 Seed storage
Sugar beet storage is not a
problem in the U.K. The cool ambient
conditions allow commercial seed to remain usable for several years. Longden & Johnson (1974b) studied the
effects of storage temperature and water content with time on seed performance
with pelleted and unpelleted Amono and Monobeet. Both higher water content and temperature lead to quicker loss of
viability over the range 5 - 18% H20 and 2 - 22°C; in the extreme
case of storage under conditions of 18% and 22°C, seeds were completely dead
after 13 months storage. Seed stored in
open containers @ 10°C produced about 9% fewer seedlings per annum relative to
the initial value (= 100) over five years, while both pelleted and unpelleted
stored in sealed thick polythene bags showed almost no decline with a water
content of 5 - 10%.
Sometimes an increase in
germination over the first year of storage was observed. This was most probably due to a combination
of saprophytic fungi on the seed surface dying and loss of post harvest
dormancy. Storing at lower temperature
(2°C) gave seedlings of lower dry weight when tested. Storage of beet seed at about 10°C is considered appropriate and
this temperature is relatively cheap and easy to maintain.
-13-
In warm humid climates seed
viability does decline and methods of minimising losses are required. Basu & Dhar (1979) found that soaking in
5 x the volume of water followed by re-drying before storage reduced the loss
of viability over three months of storage.
No chemical solution had a better effect than water but the exact mode
of action of the hydration/dehydration process is not known. There may be an effect on inhibitors in the
seed or on free radicals.
Another problem is that seed
vigour deterioration occurs before loss of viability can be determined by
standard germination tests. Seed vigour
is a measurement of the ability of seeds to germinate or emerge under
non-optimal conditions. Low vigour is
associated with slow emergence which is extremely undesirable for high sugar
yield.
2.3.3 Germination tests
The requirements for a
standard germination test for many species including Beta vulgaris are
defined by the International Seed Testing Association (Anon 1966). The conditions for sugar beet are 16 hours @
20°C, and 8 hours at 20 - 30°C per day with germination counts at 3 and 14
days. Light is not essential and the
test can be carried out on top of or between filter paper, or in sand. The seed should be pre-washed for 1 hour in
water at 25°C.
The object of the standard
test is to gain information about the field planting value of the seed under
test and to compare it with other seed lots.
The defined conditions are designed to allow the seed lot to express
regular, rapid, and complete germination, and also be repeatable within the
limits of the random sampling. However,
sugar beet field emergence is not accurately predicted by the standard test (Brown
1980).
-14-
Hibbert and Woodwark (1969)
have tried other laboratory tests using pleated paper or flat paper in sealed
containers at different temperatures and periods of counting. Results were
similar but not interchangeable and the inherent variability of seedlots makes
emergence prediction a procedure with low precision. Sand is considered to be an unsuitable
medium for germination tests but Snyder and Filban (1970) in the U.S.A. praised
a test for emergence potential of seed from a standardised sand tray.
Hibbert, Thomson and
Woodwark (1975) and Reiff (1976) cited by Johnson (1979) found that pleated
paper gave good laboratory germination, as the contact between seed and paper
was better than with flat filter paper.
Brown (1980) found low temperature results more accurate for field emergence
prediction, but this procedure is lengthy.
Bonscheur (1975) cited by Johnston (1979) found that the speed of
germination but not the final value was affected by varying temperatures and
water contents, but Heydecker, Orphanous and Chetram (1969) recommended that
care should be taken not to penalise seed by either excess or a lack of water
when under test (using garden varieties of red beet).
2.3.4 Effects of inhibitors
on germination
Many substances have been
isolated from the seed coats of sugar beet which are potentially inhibitory to
germination, most of them are organic acids.
Snyder, Sebeson & Fairley (1965) considered Oxalate to be the major
inhibitor but did note some complex interactions with others. It was concluded that effects were
specifically inhibitory rather than osmotic.
Sebeson, Mitchell & Snyder (1969) studied the effects of inhibitors
on Alpha-amylase activity on starch solutions (hydrolysis of starch is an
essential process before germination commences). The
-15-
inhibitory effects of
Caffeic, Ferulic, Gallic, P-hydroxy, Benzoic and Vanillic acids, which are
known to exist in beet seeds, were examined using Alpha-amylase and excised
embryos. Gallic acid was found to be
most inhibitory and, in general, the degree of inhibition increased with
concentration in both experiments, but the effects were less pronounced on the
embryos, indicating that some form of de-toxification must occur in the
seed. Battle & Whittington (1971)
showed that the maternal genotype influenced the early field behaviour of the
progeny through control of the level of inhibitors in the seed. In earlier experiments, Battle &
Whittington (1969b) showed that the inhibitors were situated in the perianth
and pericarp, and also that the various other acids, including Abscisic, were
involved. It was postulated that the
free phenolic acids were in equilibrium with acetone insoluble esters involved
in lignin bio-synthesis. Immature
clusters would have a relatively higher proportion of P-coumaric acid
associated with an earlier stage in lignin biosynthetic pathway.
Inhibitors may also act as 02
acceptors and affect germination in this way (Heydecker, Chetram &
Heydecker 1971).
2.3.5 The germination
process and the involvement of air and water
Water and oxygen must be
taken up before germination can occur.
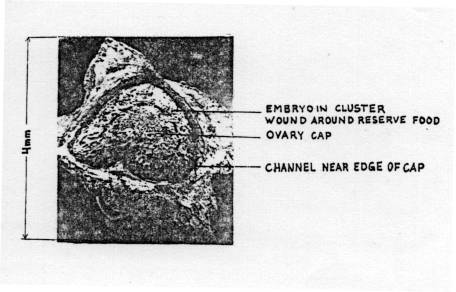
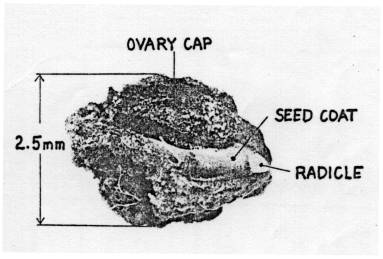
The figures on Pages 16 and 17 show that the embryo is concealed inside
the seed coat which acts as a barrier to both, but the basal pore does permit entry. The pore is not usually open, but contains
the remains of the vascular connections between the mother plant and the
embryo.
Perry and Harrison (1974)
almost completely prevented germination by blocking the pore with
Vaseline. Entry between seed cap and
the seed coat does not occur as Heydecker et al (1969) had earlier
described.
-16-
(i) Monogerm seed of variety Monohill (Aura
1975).
(ii) Opening of the seed cap and radicle emergence after two days
(Aura 1975).
-17-
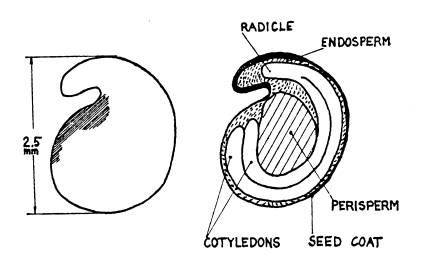
(iii) Diagrams showing external view and out-section of a true seed
(Lakon & Bulat 1958 cited by Aura 1975).
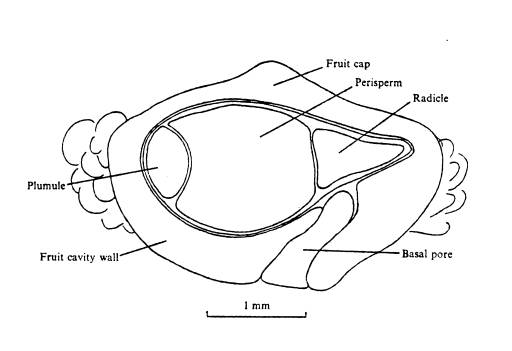
(iv) Diagram of a transverse section of a monogerm seed (Perry & Harrison 1974).
-18-
Severe inhibition has also
been described in many papers including Chetram & Heydecker (1967) and
Heydecker & Chetram (1971) by excess water in the test substrate. Excess water tends to be taken up and held
in the basal pore by capillary action.
Consequently, oxygen can only enter by diffusion, through the water at a
very slow rate.
Perry and Harrison (1974)
applied Fick's Law to the dimensions of the basal pore and estimated that, when
air filled, the oxygen diffusion flux was 4.5 ml/h and when water filled only
8.5 x 10-3 μl/h. An
embryo requires about 0.8 μl/h to germinate and therefore the process is
inhibited when the pore is full.
The observed uptake of
oxygen in a water filled pore was 0.14 μl/h. The difference from the water filled pore estimate was due to
microbial respiration.
As germination continues
cell number increases (Longden 1971) then the radical elongates forcing the
seed cap open thus making more oxygen available to the embryo facilitating
faster elongation. Coumans, Côme &
Gaspar (1976) showed that in a wet medium removing the cap before germination
resulted in more seed germinating if positioned "face up" but not if
"face down". Therefore the
external film of water could also inhibit germination. Peto (1964) cited by Heydecker and Chetram
(1971) chipped the seed cap beforehand and improved germination by both
allowing more oxygen in and reducing the mechanical effort required by the
radical to remove the cap. Heydecker
and Chetram (1971) viewed germination as more than a physiological process, i.e.
complex ecological and microbiological components are involved too. When 8 ml of water was used in a laboratory
test dish it was excessive, but adding aureomycin to inhibit bacteria leaves
more oxygen for embryos to use. Washing
seeds upsets the ecological balance between bacteria, fungi and the inhibitors
in the seed coat, and
-19-
the depressed laboratory
germination of seeds treated with fungicides may be explained by a similar
change to the micro-environment.
As germination is improved
by cap removal or inhibitor removal, whether water is excessive or not, 02
uptake and respiration must therefore precede germination and not be a
consequence of it! However, Heydecker et
al (1971) noticed that in red beet seeds when the concentration of
inhibitors was high, the oxygen uptake was high also. Therefore, some process different from normal respiration was
occurring. There may be competition for
oxygen between different metabolic pathways with imbibed seeds - inhibiting
respiration at low concentrations and uncoupling, i.e. preventing access to the
true seed, if at higher concentrations but an actual mechanism has not been
found.
Coumans et al (1976)
viewed the seed coat as a physiochemical barrier to oxygen. It (a) restricted diffusion and (b) actually
absorbed oxygen so that very little if any reached the embryo through the seed
coat.
Chetram & Heydecker
(1967) and Heydecker et al (1969) found that hydrogen peroxide in
solution was an excellent way of supplying oxygen to the embryo to improve
germination.
2.4 Factors affecting
the establishment of beet
2.4.1 Introduction
Regular sugar beet stands
used to be achieved by sowing seed at a high rate and subsequently hand hoeing
unwanted plants after emergence.
However, now that monogerm seed, precision drills and suitable
herbicides have been introduced, regular stands can be achieved without
handwork. Nevertheless, modern methods
of establishment are not always completely successful as Bray (1980)
-20-
has shown. Nearly half the area "drilled to a
stand" receives at least a small amount of handwork.
The main reason for this is
that sugar beet field emergence is difficult to predict even when laboratory
germination is known. This section
discusses the factors affecting emergence but firstly optimum or target
populations are considered.
2.4.2 Plant population
Hull and Jaggard (1971)
reviewed attempts to determine the population for maximum yield of sugar and
found many factors, viz. soil type, irrigation, sowing technique and
fertilizers, affected this optimum population.
They generally concluded that this was 65,000 plants/ha on a fertile
soil rising to 85,000 on poorer soils, but a few thousand above or below did
not seriously depress yield. Goodman
(1966) recommended 74,100/ha with a leaf area index (L.A.I.) of 2.8 (which does
not intercept all available radiation but ensures there are no non-productive
leaves). The above recommendations were
based on hand-hoed situations only where a dense crop was sown and subsequently
thinned. Draycott & Durrant (1974)
looked at populations in relation to other cultural practices, and showed
50,000 or above was adequate without hand-hoeing. It was also shown that between a rectangularity of 1:2 and 1:1
yield was not affected at higher populations (86,000/ha) but Hull & Jaggard
(1971) showed that 45 cm rows were more suitable for high populations and 60 cm
for lower populations to reduce interplant competition in both cases. Interplant competition was shown to reduce
individual plant yields considerably by Draycott & Durrant (1974) who
compared sugar yields from plants grown in a competition free plot (22,000/ha
with minimal nutrient and light competition and adequate soil water) with
denser stands.
-21-
At 22,000/ha individual
plants yielded 315 g of sugar/plant but at 81,000/ha individuals yielded only
113 g, but a much higher total yield/ha.
2.4.3 Cultivation and seed bed preparations
Cultivation for sugar beet
should make the best use of the available environmental conditions over the
preceding Autumn and Winter. The
following practices should ensure a reasonable quality seed bed in the Spring.
1)
Plough
early with a reversible plough to gain the benefits of an even surface for an
even depth of weathering, and as long a weathering period as possible.
2)
Use
as few passes as necessary with wide wheel extensions, to minimise excessive
consolidation. Use wide implements and
tandem arrangements.
3)
After
weathering use shallow cultivations only to avoid bringing clodding unweathered
material up into the seed bed.
4)
Form
a coarse tilth below the surface for drainage but a finer tilth on the surface
for water conservation.
5)
A
level seed-bed should be achieved by use of straight and rolling tined harrows
or power harrows, so that precision drills can be used and drilling depth
controlled (Spoor 1978, Clare 1976 and Rose 1972).
2.4.4 (i) Drilling
Precision drills are now
almost universal in use and are essential for the "drilling to a
stand" technique. A precision
-22-
drill is defined as one
which selects and deposits seed at predetermined distances. Common features of precision drills are:
land wheel drive, minimum seed drop, boat shaped coulters and flat rollers
(Rose 1972). In the U.K. the
"Stanhay" pinched rubber belt drill is used, but disc types are
popular elsewhere (Hull & Jaggard 1971).
Ten or twelve row machines are necessary to compensate for slow forward
speed of precision drills, but the N.I.A.E. have a test drill accurate at 11
km/hr (Hayward 1978). Munday (1977) has
shown that no commercial drill sows perfectly.
Doubles, singles or multiple misses and inconsistent spacings are always
observed, but the seed can confound the drill performance when doubles are due
to pellets with extra embryos and misses due to dead or empty ones. However, better drills that reduce the
amount of seed roll and that are more accurately space-calibrated are required
for "drilling to a stand".
Beet seeds should not be
sown below 3.8 cm due to the small perispermic reserves. Early sowing should be shallow (< 2 cm)
for good emergence. If sown later then
3 cm is better as the surface dries out (Hull & Jaggard 1971, Hibbert et
al 1975).
Alternative drilling
techniques such as fluid drilling of pre-chitted seed (Longden et al
1979, Currah 1978) have as yet unsolved technical problems and cannot be used
for fast and even emergence in sugar beet.
(ii) "Drilling to a
stand"
Drilling to a stand" is
only successful with precision drills, good emergence and relatively weed free
fields. In the U.K. 12 - 15 cm spacing
in 50 cm rows is commonly used to achieve 74,000/ha but Fletcher (1974) has
shown that no universally recommended spacing) is possible as localised factors
are involved. The technique works best
for April sowing when compensatory growth
-23-
is adequate to make up for
irregular spacing. Neeb and Winner
(1970) cited by Hull and Jaggard (1971) deliberately mixed good and dead seed
to encourage irregular spacing and reduce population and still found a linear
relation between population and yield up to 80,000/ha. However, Thomson (1956) cited by Hull and
Jaggard (1971) also deliberately obtained an irregular stand by random
hand-singling and found 0.5 t/ha less from an irregular stand than from a
similar regular, hand-singled stand.
Knott, Parker & Mundy (1976) found that with "drilling to a
stand" irregularity effects were worse with low populations, made with
wide rows and spacing, and at the same time found 70,000/ha was optimal for a
fen soil, but only 50-56,000 for a silt.
2.4.5
Field emergence
Aura (1975) categorised four
factors involved in emergence:
1) The germination energy of the seed.
2) The appearance of pathogens.
3) Mechanical soil resistance.
4) Soil, air & water content.
(i) Germination Energy.
Perry (1973) showed monogerm final emergence to be reduced by high soil
water levels and by compaction, but not by low soil temperature. However, seed lots responded differently
showing that seedling vigour is important.
(ii) Effect of fungicide.
Heydecker & Chetram (1971) showed that seed fungicide treatment
improved field performance even if laboratory tests showed the reverse.
-24-
(iii) Hegarty and Royle (1978) measured the impedance of soils
covering sugar beet seeds and showed a negative linear correlation with final
emergences. Perry (1973) also showed
how soil capping caused by irrigation water reduced emergence by 30%.
(iv) Aura (1975)
considered oxygen uptake was only seriously hindered by water in the seed or
the surrounding film, when soil water potentials were close to zero, i.e. in
very wet soils. Poor contact between
soil and seed could reduce germination by restricting water uptake in dry
soils, if the soil water potential and was less than - 10 atm no emergence
occurred. Aura also noted water
diffusion to be slower through the seed than through the soil. Pelleted seed emerges better in wet soil
conditions (Perry 1973), and Hibbert et al (1975) postulated that
pelleted seed may have a higher water requirement and should be sown slightly
deeper in dry conditions. Aura (1975),
however found pellets restricted oxygen uptake in very wet conditions.
2.4.6 Predicting sugar beet emergence
The standard germination
test is fair to the seed in that individual seeds have minimal stress and
therefore if germination is possible it should occur. However, sugar beet growers require information on field
emergence potential and therefore tests with inbuilt stresses may be more
appropriate. Perry (1973) has shown how
field stresses affect beet seed lots differentially and prior knowledge of this
would be very useful for seed selection for a particular situation.
-25-
Brown (1980) suggested low
temperature tests (5-7°C) as viable seeds which germinate at 20°C may not do so
at lower temperatures which would be experienced in the field. Longden, Johnson & Love (1970) developed
a radiography test for laboratory emergence and Longden and Johnson (1974a)
compared a radiography test with other methods, i.e. leachate conductivity, a
pleated paper test (Hibbert & Woodward 1969), and growing in compost, for
prediction of field emergence.
The radiograph prediction is
based on x-ray photographs of seed, assessed visually into "good",
"dead" and "uncertain" categories. Filled cavities, shrivelled seed, empties
and double embryos can be identified with this method. However, not all seed classified as good
will germinate, and therefore radiography tends to over-estimate laboratory
germinations. Germination in compost
was predicted equally well with radiography and standard germination
tests. For field performance the
leachate conductivity method was hopelessly inaccurate with pelleted seed and
poor with unpelleted seed and was therefore discarded as a prediction
method. The compost test was more
accurate but took three weeks and was roughly equivalent to a radiography
prediction. However, radiography does
not work for pellets. Overall the
pleated paper and the standard test gave the best prediction. It was noted that a low laboratory
germination always resulted in poor field performance, but a high laboratory
value could result in a high or low field result.
The method of prediction of
field performance was a linear regression, e.g. Y = 0.64 X + 6.2 where Y is
field emergence (%) and X is laboratory germination (%) but using a field
factor is simpler. A field factor of
72% would mean a grower could expect 72% of the laboratory germination to
emerge in the field.
-26-
The field factor can be applied
to this formula (Bleasdale 1963 cited by Longden & Johnson 1974a).
P x 100
N = ----------
G x F
Where:-
N = no. seeds required/ha
P = desired
population, plants/ha
G = laboratory
germination %
F = field
factor.
However, as sugar beet
establishment is variable, estimating the field factor may not always be
accurate enough for practical use of the formula, but it shows that improving
the field factor by preparing better seed beds will reduce the number of seeds
required to produce a regular stand.
2.4.7 The effect of seed size on emergence
Snyder & Filban (1970)
compared fruits graded 2.58 - 2.98 mm and 3.77 - 4.12 mm in diameter sown at
different depths, 3.2 cm and 5.1 cm in sand.
The deeper and smaller seed reduced establishment. The hypocotyls of the deeper sown seeds were
heavier as increased impedance forced them to widen.
In the field, emergence was
as shown:-
|
|
Depth of |
sowing |
|
Seed size (mm) |
3.2 cm |
5.1 cm |
|
2.58 - 2.98 |
37% |
20.5% |
|
3.77 - 4.12 |
51% |
38% |
Snyder and Filban (1970)
recommended seed rates should be increased by 33 per cent when using small
seed, and that small seed should always be sown at less than 2.5 cm. When soil
-27-
moisture is limiting, deep
sowing of large seed should be used. Lindsay (1980 personal communication)
showed that seed less than 2.75 mm is eliminated in processing and so only
large seed is used in practice. There is nevertheless still a size range as
investigated by Scott, Harper, Wood & Jaggard (1974). The effects of seed size on development and
yield of sugar beet were studied. small (2.8 - 3.6 mm), Medium (4.0 - 4.4 mm)
and Large (5.2- 5.6 mm) seed were used.
Overall the embryo weight
was about 20% of the "seed" and larger seeds had fewer empty
cavities. They confirmed that the large
seed within a seed lot gave a higher percentage emergence and was more
reliable.
After processing, larger
seed still performed better.
Plants produced from larger
seed had higher root: shoot ratios but at harvest plants from medium and large
seed produced similar yields. Possibly
selecting for seed size also selected different genotypes.
2.4.8 Other emergence factors
(i)
Fertilizers
Providing the recommended
rates are used (100 Kg/ha N, 100 Kg/ha P205 &
200 Kg/ha K20 MAFF 1979), fertilizers
do not have a large effect on establishment except for nitrogen which can be
used to encourage early leaf growth.
Nitrogen should be applied in the Spring at least two weeks before
sowing to allow high localised concentrations to be diluted by rainfall before
scorch damage of seedlings occurs (Last & Draycott 1979). The other nutrients, and, if required, Na
and Mg, can be applied in the Autumn (Draycott 1977).
-28-
(ii)
Fungicides
Byford (1977) studied the
effects of mercury fungicides on emergence. Maneb, Captafol and
2-(Thiocyanome-thylthio) benzothiazole (T.C.M.T.B.) seed treatments were
compared with the standard E.M.P. treatment.
No treatment was as good as E.M.P. and only maneb was similar. It has been estimated that national
emergence would be 10% lower if E.M.P. was replaced by another fungicide. Other environmental factors had much larger
effects on emergence overall, but E.M.P. treatment is definitely justified as
the untreated control had a 20% lower emergence.
(iii)
Pests
Pest control measures in
addition to seed dressing are sometimes required for pests which effect
establishment. Millipedes and beetles
can be troublesome but Gamma HCH worked into the seed bed before sowing gives
control. Wood mice which dig up seeds
and seedlings can be controlled by traps or poisoned food if numbers are high
after a mild winter (Farmers' Weekly 1979).
(iv)
Weeds
Weeds are no longer a major
problem to establishment as good herbicide control is now possible with soil
acting herbicides such as chloridazon or ethofumesate (Bray 1980).
(v) Cold injury
Cary (1975) showed that
seedlings on the point of emergence were more sensitive to frost damage than at
any stage before or after, and that no chemical used to promote winter
hardiness in other species
-29-
affected sugar beet
similarly. However, germination at low
temperature and osmotic treatments did reduce sensitivity to frost damage.
(vi) Soil temperatures
The effects of soil
temperature are more important than the actual sowing date as Scott et al
(1973) have shown. The date that the
amount of accumulated day degrees over 5°C begins to increase rapidly is a good
indicator of the onset of growing weather.
However, this can occur at any time between lst March and 20th May, but
most commonly between the lst and 10th of April. It is usually best to sow just before the onset of growing
weather.
2.5 Alternative methods of establishing beet
2.5.1 Transplanting
Scott & Bremner (1966)
investigated the potential of transplanting with multigerm varieties as an
alternative to contemporary practises and found that an extra ten tonnes/ha of
roots and more tops could be obtained.
The technique resulted in more fangy and globular roots developing, but
the desired plant populations were easily obtained. The larger leaf area duration of the transplants made them more
drought sensitive, but in the experiment drought did not seriously check
yield. High populations were not
required as it was realised that it was the period of ground cover and not the
L.A.I. obtained that had the main effect on yield.
In 1966 it was postulated
that commercial success would be possible if suitable mechanisation at an
acceptable cost could be developed but it was not forthcoming in the U.K.
However, in Japan and Bavaria transplanting is used.
-30-
More recently ADAS have
started to re-investigate the technique and preliminary trials with monogerm
varieties in paper mini-pots show promise of a 40% yield improvement (Farmers'
Weekly 1980).
Some other advantages have
been noted, namely a reduced dirt tare and better root shape which contrasts
with Scott & Bremmer, but handling costs are still prohibitive.
2.5.2 Autumn sowing
Wood & Scott (1975)
showed that October and September sowing gave full leaf cover by mid-June, thus
making more efficient use of radiation, up to 40% of the available between
April and June. Thereafter bolting
occurred and final sugar yields were never better than Spring sown crops, even
if hand rogued. Ethephon applied at 10 g/1 partly controlled bolting but killed
55% of the plants. At a lower dose (2
g/1) it was ineffective.
It was concluded that Autumn
sowing was unsuitable for the U.K. unless extremely bolting resistant varieties
could be developed. This is unlikely as
flowers are needed for breeding.
Autumn sowing is also
inadvisable as virus yellows and downy mildew would be carried over more
easily. Autumn sowing is practised in
warmer climates, viz. Italy and Japan
where low temperature induced bolting is not a problem.
2.5.3 Conclusion
In this review, sugar beet
establishment has been shown to be the "weak link" in the production
of high sugar yields. The various
alternatives for all stages from seed production to establishment all have too
many drawbacks to use in practice. The
following experimental section is principally concerned with methods of testing
and improving the laboratory performance of beet seed, which may result in
better field establishment and higher yields.
-31-
3. EXPERIMENTS
3.1 Materials and Methods
3.1.1 Description of Seed Lots
The general objective of the
experimental work was to compare the germination performance of five different
lots of sugar beet seed under several different test conditions with various
treatments, and relate the performance to features of the seed lots. The five seed lots used were as follows:-
Lot I, variety Monotri,
harvested in 1977;
Lot 2, variety Monotri,
harvested in 1975;
Lot 3, Bush Mono
'"G", harvested in 1979;
Lot 4, Amono, harvested in
1979;
and Lot 5, Nomo, harvested
in 1975.
Lots 1 2 and 5 had been
lightly rubbed after harvesting but not further treated, while Lots 3 and 4 had
received the complete commercial processing and pelleting procedure as
described in Sections 2.2.6 and 2.2.7.
3.1.2
Seed Characteristics
One hundred air-dried seeds
from each lot were weighed, (before and after de-pelleting by washing in
running tap water where necessary). The
true seeds were dissected out from all seeds by lifting the seed cap with a
dissecting needle, after soaking in water for 12 hours, and then weighed individually. The relationships between total and true
seed weights were determined. Moisture
contents of each seed lot were also determined using an infra-red moisture
meter and milled seed samples.
-32-
3.1.3 Germination and Emergence Tests
The purpose of Experiment 1
was to test all five seedlots under conditions similar to those defined by the
International Seed Testing Association (ANON 1966), as described in Section
2.3.3.
Germination counts were
taken at intervals to determine the final germination percentages, and mean
germination time (M.G.T.).
The procedure used for this test was as follows:-
1. Four groups of fifty seeds were counted out from each of the
rubbed seed lots. Small, damaged and
multigerm seeds were excluded as it was assumed further processing would also
have removed them.
2. Four groups of fifty pelleted seeds were counted out from the
commercial seed lots (without selection).
The pelleting material was removed by washing.
3. All seeds were soaked in tap water for approximately l½ hours.
4. After a short period of air drying, each lot was placed in a 9
cm petri-dish containing three Whatmans' grade 181 filter papers and 5 ml of
distilled water. The dishes were
covered and placed in a temperature controlled incubator without
illumination. The temperature was maintained
at 20°C for 16 hours and 25°C for 8 hours per day (standard temperatures).
5. In this test germination counts were made initially at 2 day
intervals, but the counting interval increased as germination approached
completion.
6. Seeds were counted as having germinated when the radicle had
forced the seed cap open and could be seen emerging from it.
-33-
7. As some of the selected seeds contained more than one true
seed, despite the attempt to exclude them, they were considered to have
germinated if one or more radicles appeared.
This procedure was chosen
after considering the observations of
Chetram and Heydecker (1967), Perry and Harrison (1974), and Hibbert and
Woodwark (1969). Five ml of distilled
water on three filter papers was known not to be excessive for
germination. Experiment 1 was repeated
at the end of the experimental period, (October 1980 - February 1981) to test
for changes in germination performance in any of the seed lots.
The aim of Experiment 2 was
to test seed germination at a lower temperature than the standard
recommendation. It was suggested by
Brown (1980) that spring-sown seed experiences seed bed temperatures well below
those recommended for the standard test (ANON 1966). The procedure used was the same as in Experiment 1 except that
the incubator was maintained at a constant 7.5 C, and also seed lots 2 & 5
were started two days before the others.
This was because it was anticipated that these lots would take longer to
reach the period when most seeds germinate.
The temperature was selected to be low enough to allow germination, but
not so low as to considerably prolong the duration of the experiment (Brown
1980).
The petri-dishes used in
Experiment 2 which still contained ungerminated seeds after 30 or 32 days for
Lots 1, 3 & 4, and 2 & 5 respectively were transferred to a cabinet at
standard temperatures as germination at 7.5°C was considered to have been
complete.
Germination counts were
taken to assess the proportion of seeds which would germinate under standard
conditions but not at 7.5°C. After a
further 19 days germination at standard temperatures was considered complete.
-34-
The remaining ungerminated
seeds were dissected to determine qualitatively if the true seeds were
shrivelled or absent, or apparently normal.
Experiment 3 was a repeat of
both Experiments 1 and 2 with advanced seed.
The aim was to test the effect of advancing as a seed treatment, on
germination performance at both standard and low temperatures. The advancing procedure used was as follows
(Longden 1971):-
1. Samples from the pelleted seed lots were washed to remove the
pelleting material.
2. Approximately 1000 seeds from each of the rubbed seed lots
were weighed and placed in a 9 cm petri-dish.
3. Stage 2 was repeated with the de-pelleted seeds.
4. Tap water was added to each dish in an amount approximately
equal to the weight of each seed sample in each dish.
5. The dishes were covered for 24 hours at room temperature, then
uncovered with the contents spread out to dry for a further 48 hours.
6. Stage 5 was repeated twice, so that each seed sample received
a total of three advancing cycles.
The seeds were then counted
and set up as for Experiments 1 & 2, except that the 1½ hour pre-soak was
omitted. The test at standard
temperature was named Experiment 3 (i), and the low temperature test Experiment
3 (ii). It was noticed that during the
advancing procedure some seeds germinated, and seed caps became detached from
others, particularly in the de-pelleted samples. This was also observed by Longden (1971, 1973). However, only seeds entirely intact after
the advancing treatment were selected for testing, and in Experiment 3 (i) seed
Lots 3 & 4
-35-
were not used as there were
too few intact seed left in the advanced seed stocks.
The aim of Experiment 4 was
to test germination performance with a solution of Gibberellic acid (GA3)
in place of distilled water. The
procedure used was therefore the same as in Experiment 1 except that 5 ml of a
100 ppm solution of GA3 was placed in the petri-dishes, and
germination counts were initially made at daily intervals. The concentration used had earlier been
found to be optimal for improving germination performance if used in a 24 hour
steep before testing (Scott et al 1972).
Experiment 5 was a repeat of
Experiment 3 (i) (advancing and testing at standard temperature) except that,
the tap water used for advancing was replaced by a 100 ppm GA3 solution. The aim of this experiment was to test the
effects of advancing with GA3 on germination performance. As Experiment 5 was also effectively a
treatment combination test, either additive effects or interactions may be
observed.
The final test (Experiment
6) was an emergence test. Four
replicates of 100 seeds or pellets from each seed lot were sown 2 cm deep in
trays containing John Innes number 3 compost.
The trays were placed in an illuminated (16 hour photo-period) glass
house at approximately 16°C. The aim of
this experiment was to examine the relationship between germination and
emergence from compost, of the seed lots.
After 16 days seedlings were
counted and cut off at soil level. The
dry weights of the cut seedlings were assessed after oven drying for 24 hours
at 90°C. A final count was made 27 days
after sowing for slower emerging seedlings but no dry weights were recorded.
-36-
3.1.4 The effect of water-soluble seed extracts on cress seed
germination
1. 30 g of seeds from Lots 1, 2 & 5 were pulverised in a hand
mill.
2. The milled sample obtained was mixed with 80 ml of distilled
water, periodically shaken and left in sealed bottles for 2 days.
3. Five ml of liquid extract was pipetted into petri-dishes
containing 3 Whatmans' grade 181 filter papers and 50 cress (Lepidium
sativum) seeds. The dishes were
incubated as in Experiment 1 with daily counts.
4. Stage 3 was repeated with the extract diluted to 0.5, 0.25,
0.1 and 0.01 of the original concentration.
Cress seeds were counted as having germinated when radicles >1 mm
were observed. The final germination
percentages and M.G.T. of the cress seeds were determined. Lots 3 & 4 were not used for this
determination as chemical treatments in the processing procedure may have
interfered with cress germination.
3.2 Results
3.2.1 Seed Characteristics
The mean values for various
seed characteristics are presented in Table 1. A large proportion of the pelleted seed lots was in the form of
the clay pelleting material, but after de-pelleting the mean seed weights of
the pelleted seed lots (Lots 3 & 4) were similar to the mean weights of Lots
1 & 5. However, the mean seed
weight of Lot 2 was lower than the others.
-37-
TABLE 1: SEED
CHARACTERISTICS
|
SEED |
|
|
SEED LOT |
|
|
|
CHARACTERISTICS |
1 |
2 |
3 |
4 |
5 |
|
Mean Pelleted weight (mg) |
- |
- |
69.4 |
69.1 |
- |
|
Mean seed weight (mg) |
10.39 |
8.67 |
11.24 |
11.57 |
11.29 |
|
Standard Deviation |
±1.62 |
±2.10 |
±2.11 |
±2.41 |
±1.37 |
|
Co-efficient of variation |
±15.6 |
±24.2 |
±18.8 |
±20.8 |
±13.2 |
|
Proportion of pellet as seed (%) |
- |
- |
16 |
17 |
- |
|
Moisture content of seed (%) |
10.4 |
10.2 |
9.4 |
9.2 |
9.8 |
|
Mean true seed weight (mg) |
3.73 |
2.92 |
4.66 |
3.8 |
3.46 |
|
Standard Deviation |
±1.3 |
±1.13 |
±1.10 |
±1.14 |
±0.87 |
|
Co-efficient of variation |
±34.9 |
±38.7 |
±23.3 |
±30.1 |
±24.9 |
|
Mean proportion of seed as true seed |
36.5 |
33.6 |
41.9 |
32.3 |
30.1 |
|
Standard Deviation |
±13.45 |
±11.65 |
±8.13 |
±9.08 |
±7.49 |
-38-
The mean true seed weights
of Lots 1, 4 & 5 were similar, while Lots 2 and 3 were lower and higher
than the other lots. The mean
proportion of the total seed weight as true seed weight was similar for all the
seed lots except Lot 3 which had a greater proportion than the others. The moisture contents of the five seed lots
were also similar.
The relationships between
seeds and true seed weights are presented in Figures 1-5. Integers plotted on the figures indicate the
number of seeds with the same seed and true seed weights.
The relationships revealed
highly significant correlations (P = 0.001) with Lots 2, 3 & 4, a good
correlation with Lot 5 (P = 0.01) but a non-significant correlation with Lot 1.
The slopes of the
regressions are slightly greater than 1 for Lots 2, 3 & 4 but much less
than 1 for Lots 1 & 5. There would
appear to be an association between the correlation coefficient and the slopes
of the regressions. However when the
actual true seed weights and mean proportions of the seed weight as true seed weight
are compared Lots 1 and 5 have the largest and smallest standard deviations in
both cases (Table 1).
The coefficients of
variation for the true seed weights of all 5 lots are all larger than the
respective coefficients for the total seed weights indicating that there is
relatively more variation in the true seed weights.
3.2.2 The Germination Experiments
The germination patterns for
the six germination experiments are presented in Figures 6-11. The final germination percentages and mean
-39-
FIGURE 1: THE RELATIONSHIP BETWEEN SEED WEIGHT AND
TRUE SEED WEIGHT FOR SEEDLOT 1. LINEAR
REGRESSION: Y = 0.218 X +9.58 WHERE Y
IS SEED WEIGHT AND X IS TRUE SEED WEIGHT. CORRELATION: 0.1756 (NOT SIGNIFICANT)
-40-
FIGURE 2: THE RELATIONSHIP BETWEEN SEED WEIGHT AND
TRUE SEED WEIGHT FOR SEEDLOT 2. LINEAR
REGRESSION: Y = 1.09 X +5.49 WHERE Y IS
SEED WEIGHT AND X IS TRUE SEED WEIGHT. CORRELATION: 0.5872 (SIGNIFICANT AT P
= 0.001)
-41-
FIGURE 3: THE RELATIONSHIP BETWEEN SEED WEIGHT AND
TRUE SEED WEIGHT FOR SEEDLOT 3. LINEAR
REGRESSION: Y = 1.19 X +5.72 WHERE Y IS
SEED WEIGHT AND X IS TRUE SEED WEIGHT. CORRELATION: 0.6101 (SIGNIFICANT AT P
= 0.001)
-42-
FIGURE 4: THE RELATIONSHIP BETWEEN SEED WEIGHT AND
TRUE SEED WEIGHT FOR SEEDLOT 4. LINEAR
REGRESSION: Y = 1.06 X +7.55 WHERE Y IS
SEED WEIGHT AND X IS TRUE SEED WEIGHT. CORRELATION: 0.502 (SIGNIFICANT AT P =
0.001)
-43-
FIGURE 5: THE RELATIONSHIP BETWEEN SEED WEIGHT AND
TRUE SEED WEIGHT FOR SEEDLOT 4. LINEAR
REGRESSION: Y = 0.521 X +9.48 WHERE Y
IS SEED WEIGHT AND X IS TRUE SEED WEIGHT. CORRELATION: 0.329
(SIGNIFICANT AT P = 0.01)
-44-
germination times (M.G.T.)
with least significant differences at P ³0.05 are presented in Tables
2 & 3. The formula used to
calculate the M.G.T. for individual replicates was:-
Σ(G x T)
(M.G.T.) = --------------
F
Where T = the
day on which germination count was made
G = the
number of seeds germinated on the day of the count
F = final
number of seeds which germinated in each replicate
This formula was used in all
germination experiments (Battle & Whittington 1969a).
All significant differences
referred to in this section and thereafter unless otherwise indicated are at
significance level P ³0.05.
The six germination tests
were carried out over a period of time and therefore direct comparison of seed
lots across experiments is not valid.
Statistical analyses were only carried out within each germination test.
The first germination test
(Experiment 1) carried out under standard (i.e. optimal) conditions for
germination showed that seed lots 1, 2, 3 & 4 had almost completed
germination after four days, while Lot 5 had an extended germination period (Figure
6). Lots 1, 3 & 4 reached
higher final germination percentages than Lots 2 & 5 (Table 2). Lot 2 was significantly lower than Lots 1, 3
& 4 and Lot 5 was significantly lower than Lot 2.
The most rapid germination
occurred with Lots 3 & 4 which were not significantly different from each
other (Table 3). Lots 1 2 &
5 however,
-45-
TABLE 2: FINAL GERMINATION
PERCENTAGES IN
SIX EXPERIMENTS
KEY P = 0.05 *, P = 0.01
**, P = 0.001 ***
SIGNIFICANCE LEVEL SL,
LEAST SIGNIFICANT DIFFERENCE
LSD,
STANDARD ERROR DIFFERENCE
SED.
|
EXPERIMENT |
|
|
SEED LOT |
|
SED |
LSD @ |
SL |
|||
|
|
1 |
2 |
3 |
4 |
5 |
|
P = 0.05 |
|
||
|
1. Standard test |
97.5 |
84.0 |
93.0 |
95.0 |
65.0 |
2.63 |
5.72 |
*** |
||
|
2. Constant 7.5°C |
82.5 |
63.5 |
82.5 |
81.0 |
61.0 |
5.24 |
11.43 |
*** |
||
|
3(i). Advanced, standard temperatures |
95.0 |
88.5 |
- |
- |
82.0 |
2.74 |
7.75 |
** |
||
|
3(ii). Advanced, constant 7.5°C. |
93.5 |
78.5 |
95.0 |
92.5 |
76.0 |
4.89 |
10.65 |
** |
||
|
4. GA3 in petri-dish |
93.0 |
83.0 |
96.5 |
97.5 |
79.0 |
2.49 |
5.42 |
*** |
||
|
5. GA3 Advanced,
standard temperatures |
95.5 |
88.0 |
- |
- |
81.0 |
4.10 |
10.01 |
* |
||
